
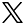

数理モデルでつなげる脳の仕組み vol.2
汎用性のある臨界期のモデル
この理論モデルについて、概略を説明しましょう。まず、抑制性ニューロンの影響が弱い臨界期前の段階では、一次視覚野のニューロンは視覚刺激による入力に加えて、常に両方の目からの経路に沿って生じる自発的活動も入力として受け取っていると考えます。この状況では、たとえ視覚刺激が片方の目に偏っていたとしても、自発的活動のために、左右の目からの経路の入力に大きな偏りは生じず、したがってシナプス可塑性によって左右の目のバランスが大きく崩れることもないという結果になります(図2)。
外からの視覚刺激に対する自発的な発火の割合を、マウスによる実験で調査。臨界期が始まると、視覚刺激(外部からの情報)の影響が大きくなる。
しかし、抑制性ニューロンの影響が強くなる臨界期では、ニューロンは強い抑制を受けるため、自発的発火による入力だけでは十分に発火できず、強い視覚刺激を受けたときにしか発火しなくなります。この状態で片方の目を塞ぐと、左右の目からの入力に対するニューロンの応答に大きな偏りが生じ、そのような状態で起こるシナプス可塑性の結果として、ニューロンは開いている目からの情報を優先して処理するようになると考えたのです。
単純なモデルですが、この理論でいくつかの実験結果を説明できます。たとえば、臨界期の前後でどのくらい細胞の応答性が変化するかなどです。そうした他の実験の説明ができることで、内的情報と外的情報による神経回路の学習の変化と、臨界期の開始が関連しているかもしれない、という理論は正しそうだと思いました。
そこで、理論ではなく実験が専門の研究者たちと組んで、臨界期が始まるときに、本当に自発的な発火の影響が変わっているのかを調べてもらいました。すると、その理論で予測したことと同じような結果が得られたのです。
臨界期にも、さまざまな種類があります。たとえば聴覚なら絶対音感、あるいは言語のリスニング能力などがそうです。
個々の臨界期に応じた脳の領野で、たとえば抑制性が強くなり、自発的な発火から外的な入力による発火へと切り替わって学習が進む。そして、その学習の信号が切り替わる時期が早いか遅いかに応じて、脳に低次から高次への階層的な構造ができるのではないか。それが臨界期であるというのが、私の解釈です。
この臨界期の理論は、従来のものより幅広い期間の学習を、発達段階によらず一つの法則で説明することができました。
臨界期は、さまざまな神経回路にあります。どこまで神経回路のメカニズムが解明されているのかは脳の場所次第なのですが、たとえば、聴覚や視覚といった大脳新皮質の感覚野では、かなり研究が進んでいます。私の作った理論は、学習信号の内部由来から外部由来への切り替えという抽象的性質に焦点を当てることで、個々の脳領域の差異によらず、より一般の臨界期現象に適用できる可能性があります。そうした理論からの提案によって、より理解が進むことがあると思うのです。
暴走を止めているのは誰?―「ヘブ則」安定化のミステリー
もう一つ別の研究を紹介します。神経回路の学習における基本法則についての研究です。この基本法則は古くから提唱されていて、カナダの心理学者ドナルド・ヘッブが1949年に自著に書いた「ヘブ則」という仮説がもっとも有名です。定性的には一部の実験結果と整合性があるので、広く信じられている仮説なのですが、じつはヘブ則をそのままモデルに入れると、いろいろ困ったことが起きるのです。
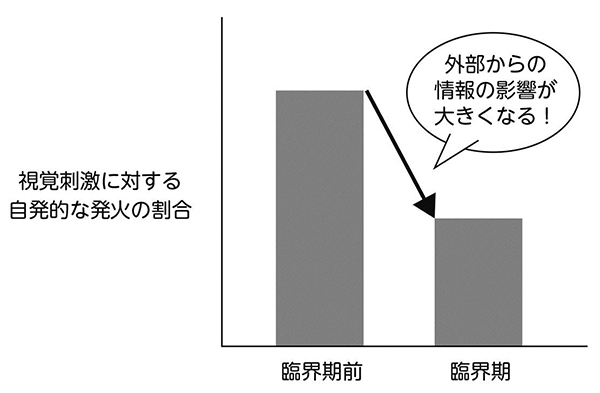
たとえばヘブ則では、つながっているニューロン同士が一緒に発火すると、ニューロン同士をつなぐシナプス強度が強くなります。図3はヘブ則がうまく働いたときの連想記憶のモデルですが、このようにうまくいくケースばかりではありません。じつはこれは、すごく不安定な状態なのです。
ある人の顔と声をそれぞれ符号化しているニューロン群がある。その人と会って話をすると、二つのニューロン群が一緒に発火し、それらをつなぐシナプス強度が強くなる(ヘブ則)。その結果、次回から電話の声だけで相手の顔を連想するようになる(連想記憶)という考え方。
ある程度、強固につながっているニューロンのグループがあると、集団として発火が強くなってしまい、周囲にあるニューロンまで巻き込んで発火させてしまうのです。どんどん周囲に侵食して、新しいメンバーをグループに引き込み、ひたすらグループの規模を拡大させていきます。そうしたことが、ヘブ則を入れたモデルでは確認できます。これは、とても困ったことです。
どんな困ったことが起きるのか、例を挙げてみましょう。ある記憶に関わる神経回路の中で、あるニューロンのグループが昨日の晩ご飯のおかずを符号化していて、別のグループが今日の昼ごはんを符号化しているとします。そのとき、昼ごはんを符号化したグループがより大きく活動すると、晩ごはんのグループは相対的に小さくなって昼ごはんのグループに侵食されて、晩ごはんの記憶が薄れてしまいます。この昼ごはんのグループはさらに大きくなっていって、食べ物といったら今日の昼ごはんのことしか考えられなくなってしまうかもしれません。
シナプス可塑性の法則として、ヘブ則はいろいろな実験データとも整合するのですが、それだけで神経回路の発達や学習過程を説明するのは、難しいようです。そこに根本的な、あるいは理論的なミステリーがあります。脳の中で、何が、どのようにして、ヘブ則の暴走を安定化させているのでしょうか?
謎を解くためには、さまざまなアプローチがあり、これは私が現在も研究しているテーマの一つでもあります。ここで紹介するのは、ヘブ則とは別の可塑性の法則の存在です。その法則とは、神経回路の発火の恒常性に関する可塑性です。
恒常性とは、変化を最小限に留めようとする性質です。つまり、神経回路の恒常性とは、回路が興奮しすぎたら抑制し、抑制されすぎていたら興奮させるのです。
調べたのは、複数ある恒常性の可塑性の一つで、神経回路の活動全体を上下させるメカニズムです(図4)。
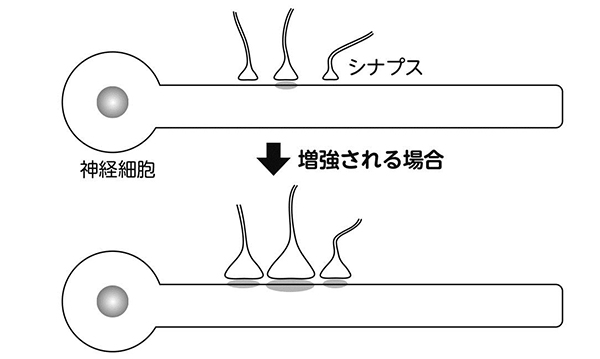
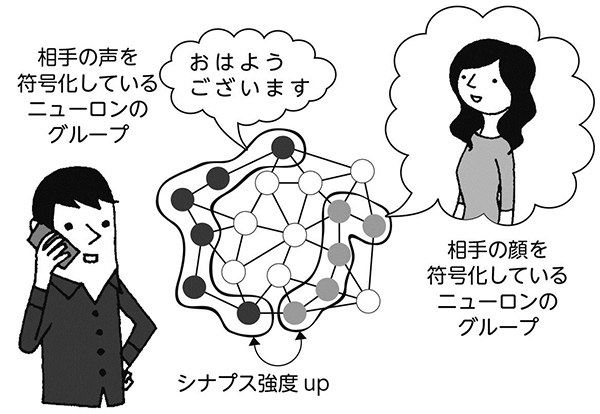
各シナプスからの相対的な入力の強弱を変えずに、全体を調整するメカニズム。図ではシナプス全体の強度を強めた場合を示している。
ヘブ則は、個々のシナプス強度を調整する細かなメカニズムですが、ある種の恒常性の可塑性は、複数あるシナプス全体の強度を大きくしたり、小さくしたりします。たとえるなら、画像のコントラストを上げたり下げたりするようなものでしょうか。
強いシナプスも弱いシナプスもある状態で、強いものはもっと強くなり、弱いものもそれなりに強くするようなメカニズムが、神経回路の可塑性にあるのです。
この2種類の可塑性を組み込んだモデルは、これまでにもいくつか提案されていましたが、それらの時間スケールの違いという重要な特徴を考慮に入れていませんでした。
その理由は、最近になってようやく、この恒常性の可塑性を担っている遺伝子が報告され、その遺伝子改変マウスを使ってヘブ則の効果と恒常性の可塑性の効果を切り分けて観測できるようになったからです。
私が研究を始めた頃、視覚野で行われた実験で、二つの可塑性(ヘブ則と恒常性の可塑性)は時間スケールがまったく違うということが発見されました。実験的に片目を閉じさせてから、その状況を学習する過程を調べるわけです。
ヘブ則の可塑性は、目を閉じて3日後には大きな効果が出ます。閉眼側からの入力が、どんどん弱くなるのですね。
ところが、恒常性の可塑性の開始は、目を閉じてから3日経ってもまだ始まりません(図5)。つまり実験的には、まず閉眼側の入力が弱まり(ヘブ則)、それより遅れて両眼の入力が強まる(恒常性の可塑性)ことが分かったのです。
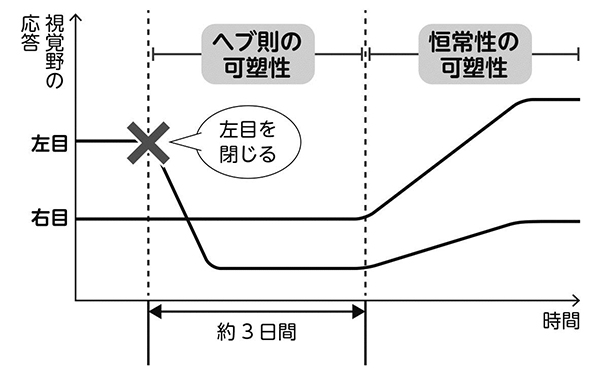
左目を閉じると視覚野の応答が下がり始め(ヘブ則の可塑性)、それを補うように3日後から両目の視覚野の応答が上がる。(恒常性の可塑性)。
著者:豊泉太郎 神経適応理論研究チーム チームリーダー
出典:講談社ブルーバックス
つながる脳科学(数理モデルでつなげる脳の仕組み) もくじ
- 数理モデルでつなげる脳の仕組み vol.1
- 数理モデルでつなげる脳の仕組み vol.2
- 数理モデルでつなげる脳の仕組み vol.3
- 数理モデルでつなげる脳の仕組み vol.4
- 数理モデルでつなげる脳の仕組み vol.5